






 Author Affiliations
Author Affiliations
Abstract
Inflammatory bowel disease (IBD) is a multifactorial disease of the digestive tract. Their prevalence has increased considerably over the previous decade due to industrialization and changes in people’s lifestyles. These are incurable diseases whose management is not unanimous because patients react differently to the treatment administered. Therefore, the search for new therapeutic targets is continuous to find better treatment, to reduce the periods of crisis, and to improve the health of the patients. In view of this fact, we have in this article brought together the various therapeutic targets associated with these pathologies to produce effective care against inflammatory bowel diseases.
Keywords
Inflammatory bowel disease, Crohn’s disease, Ulcerative colitis, Cytokines, Shingosine-1-phosphate, Exosomes, Transcription factors.
Introduction
Inflammatory bowel diseases are chronic and recurrent conditions affecting the digestive tract.[1] They are represented by various conditions such as ulcerative colitis, crohn’s disease, and undetermined colitis. Crohn’s disease differs from other IBD in that it is located in the entire gastrointestinal tract, with an initial location in the ileum and cecum, whereas ulcerative colitis is mainly found in the colon.[2] The main manifestations of these conditions are non-bloody diarrhea, abdominal pain, and the appearance of a yellowish fatty mass in the lower right quadrant of the abdomen.[3] Fever and myasthenia with complications such as fistulas to the bladder, hepatitis abscesses, lymphatic aggregation and growth retardation in children. The expansion of these diseases across the globe is a result of industrial advances and the lifestyle of populations. Indeed, the highest incidence of Ulcerative colitis (UC) has been currently recorded in Europe (24.3/100,000), while the new cases of Crohn’s disease (CD) have been mainly recognized in North America (20.2/100,000).[4] Although all ages are predisposed to the disease, there is a high incidence between 17-39 years for Crohn’s disease and 40-64 years for ulcerative colitis. Females are 2.83% more likely to be affected by these diseases than males. However, the spread of these diseases around the world is increasing. Indeed, numerous cases in Asia, Latin America, and Africa show the need to take repressive measures to stop their spread.[5]
Although their etiology is poorly understood, a multifactorial cause has been stated namely genetic predisposition involving the expression of 242 genes including autophagy related 16 like 1 (ATG16L1), cadherin 1 (commonly known as E-cadherin; CDH1), hepatocyte nuclear factor 4 alpha (HNF4a), interleukin 10 (IL10), interleukin 10 receptor alpha (IL10RA), interleukin 10 receptor beta (IL10RB), interleukin 23 receptor (IL23R), immunity-related GTPase M (IRGM), leucine-rich repeat kinase 2 (LRRK2), nucleotide-binding oligomerization domain containing 2 (also known as CARD15; NOD2), and protein tyrosine phosphatase non-receptor type 2 (PTPN2), which are involved in innate mucosal defense, Paneth cells, epithelial barrier, immune cell recruitment, antigen presentation, T cell regulation, immune tolerance, endoplasmic reticulum stress, cellular oxidative stress and cellular apoptosis.[6-12] Environmental factors such as tobacco, food, drugs, and immune dysfunction. Irritation of the intestinal wall by food, certain xenobiotics, or bacteria from the microbiota results in elevated expression of toll-like receptors (TLRS 4 and 5) by the cells of the intestinal wall (enterocytes, paneth cells, etc.), which promotes abnormal recognition of antigenic markers.[12] There is a continuous recruitment of macrophages, neutrophils, and dendritic cells that produce pro-inflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-12, and IL-2 following the activation of nuclear factor (NF-kB). There is also the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) by macrophages and excessive production of lysosomal myeloperoxidase (MPO) by neutrophils and macrophages, which destabilize cell membranes. This process is maintained by TH1 and TH17, which produce IFN-γ, IL-17, TNF-α, and IL-12, promoting differentiation into the M1 phenotype of macrophages at the expense of the M2 phenotype.[13-15] There is also an inhibition of regulatory T cells (Tregs).[1] Similarly, inhibition of the production of hemoxygenase 1 (HOX-1) by inhibition of the NRF-2 factor results in a decrease in carbon monoxide (CO), which, in normal conditions, prevents the function of antigen presentation by dendritic cells and macrophages and allows the activation of Tregs.[16-17]
IBD is a disease that is currently incurable and for which the search for new therapeutic targets is ongoing. Medical research aims to increase the periods of remission and decrease the periods of crisis, even though with age, the periods of remission become shorter and shorter. As a result, several therapeutic targets in the signaling pathways of IBD have been identified for effective treatment, although the search for new therapeutic targets is ongoing.
Methodology
For this review study on therapeutic targets associated with the treatment of IBD, the databases PubMed and Embase were systematically studied. Publications on studies of IBD, mainly on CD and UC, were selected. Subsequent screening was performed. This allowed only articles on the study of specific markers of IBD to be retained. Therefore, a list of therapeutic targets was established during drug research associated with IBD. Articles that did not address IBD were excluded, as well as those that were not in English or French.
Results & Discussion
Various published articles were reviewed, and six groups of therapeutic targets were referenced.
Integrins: These are heterometric transmembrane receptors of leukocytes formed by 2 subunits, α and β. The α4β7 integrin are found on gut-related lymphocytes, α4β1 are present on the surface of leukocytes and αEβ7 (Table 1) are expressed by intraepithelial T cells, dendritic cells and regulatory T cells, On vascular endothelial cell adhesion molecule-1 (VCAM-1), mucosal addressing cell adhesion molecule-1 (MadCAM-1) of endothelial cells and E-cadherin on mucosal epithelial cells.[18-19] During intestinal inflammation, there is an increase in the expression of cell adhesion molecules that induce the recruitment of leukocytes and lymphocytes to the intestinal mucosa, which are essential for the maintenance of inflammation.[20]
| Targets | Roles | Antagonists | |
| integrins | α4β7 | Allows adhesion of CD4+ LT to the intestinal wall.[19] |
Natalizumab, Vedolizumab |
| α4β1 | Allows leukocytes to adhere to endothelial cells.[19] |
Natalizumab | |
| αEβ7 | Allows adhesion of TH9 and CD8+ LT to the intestinal mucosa.[19] |
Etrolizumab | |
| Cytokines | tnf-α | Recruitment of inflammatory cells, induction of pro-inflammatory cytokine production.[24] |
Infliximab, Adalimumab, Golimumab , Certolizumab |
| IL-12 | Induces differentiation of TH to TH1 in the presence of activated CD4+.[25] |
Ustekinumab, Risankizumab, mirikizumab | |
| IL-23 | Promoting the training of TH17.[26] | ||
| Receptors | Sphingosine-1 Phosphate receptor | Trigger the signal that led to recruitment of immune cells by the binding sphingosine-1 phosphate.[19] | Ozanimod, Etrasimod |
Table 1: Some therapeutic targets and their antagonists
Cytokines
TNF-α: It is an essential cytokine for the maintenance of the inflammatory response. It is involved in various biological activities such as apoptosis, cell proliferation, inflammation, and many others.[1,21] Overexpression of this cytokine is the cause of several chronic diseases such as IBD, rheumatoid arthritis, and psoriasis. Poor regulation of the metabolism of this cytokine leads to the maintenance of a permanent inflammatory state (Table 1). It binds two specific receptors: TNFR1, which is expressed by all human cells and is therefore a key point in TNF-α signaling, and TNFR2, which is present on immune cells.[22,23] TNFR1 signaling is involved in cytotoxic and proinflammatory responses, while TNFR2 signaling is involved in cell proliferation, activation, and migration. It is the main therapeutic target involved in drug discovery for IBD.[24-26]
IL-12 AND IL-23: These are pro-inflammatory cytokines involved in the pathogenesis of IBD. IL-12 consists of 2 subunits p35 and p40 while IL-23 consists of p19 and p40 (Table 1). IL-12 is involved in the activation of CD4+ lymphocytes and the differentiation of T cells into TH1, which are producers of interferon (IFN) γ, which is a proinflammatory cytokine. IL-23 is involved in the formation of TH-17, which also produces IFN-γ and IL-17.[27-30]
Enzymes
Janus Kinase: It is a family of 4 tyrosine kinases (JAK1, JAK2, JAK3, and tyrosine kinase 2) that interact with a family of 7 proteins: The Signal Transducers and Activators of Transcription (STATs).[31] JAK3 is expressed specifically by TH-17, while the other 3 are ubiquitous.[32] JAK2 is involved in IL-23 production, while TYK2 is involved in IL-12 production. Those two interleukins are the most implicated in Crohn’s disease physiopathology. The JAK-STAT signaling pathway is involved in the production of 60 cytokines, hormones, and growth factors in Crohn’s disease and IL-5, IL-9, IL-13, and IL-33 for ulcerative colitis (Table 2). This signaling pathway plays an important role in innate and adaptive immunity, hematopoiesis, cell growth, and cell differentiation.[33-36]
| Enzymes | Products | Roles | Sources | Inhibitors |
| JAKinase | When activated by cytokines binding, Phosphorylate STATs which dimerize and fix in DNA to initiate proinflammatory cytokines genes transcription. | TH17 for JAK 3, and all cells for others JAKs | Tofacitinib, Upadacitinib | |
| myeloperoxidase | HClO | Produce HClO from H2O2, which bind to cells cytoplasmic membranes and maintain oxidative stress in IBD. | Most neutrophiles ;
Few macrophages |
|
| NADPH oxydase | O2. | Produces superoxide anion which bind to cells cytoplasmic membranes and maintain oxidative stress in IBD. | Most neutrophiles ;
Few macrophages |
|
| Hemoxygenase-1 | CO ; Fe ; Biliverdine | CO inhibits phagocytose and activates Tregs, Biliverdine inhibits activation of genes imply in productions of COX-2, NO synthase. | Enterocytes, macrophages | tin-protoporphyrin (SnPP) |
| Phosphodiesterase-4 | AMP | Degradation of cAMP | Most in immunocytes | Apremilast |
| Cyclooxygenase-2 | PGE2, PGI2 | Produces prostaglandins that involved in inflammatory process for homeostasis. | Inductible in Alls mammalian cells during inflammatory status | Licofelone, naproxen |
| 5-Lipoxygenase | Leukotrien, Lipoxins | Produces Leukotriens and Lipoxins that are involved in inflammatory process for homeostasis. | Zileuton, MK-866 |
Table 2: Implications of some enzymes in inflammatory bowel disease and their inhibitors
Myeloperoxidase: It is a dimer present in high concentration (2 to 5% by weight) in the primary granules of neutrophils and in small quantities in monocytes, which lose it during their differentiation into macrophages.[37-39] It is involved in many pathologies such as arthritis, lung infections, and IBD. The neutrophils responsible for their production have a strong affinity with the intestinal endothelium. This enzyme is released into the intestinal lumen during inflammation during neutrophil degranulation.[40] It catalyzes the hydrolysis reaction of H2O2 to HClO, which oxidizes the intestinal cell membranes (Figure 1). During inflammatory bowel diseases, there is an increase in the production of this enzyme, which is correlated with the level of neutrophils present in the intestinal lumen (Table 2). It plays an important role in cellular messenger immunity via the activation of transcription factors (NF-κB) and in the induction of apoptosis via the caspase pathway. Its production by macrophages remains limited. Similarly, the HClO produced activates the nuclear translocation of the NF-κB factor, which initiates the transcription and synthesis of many pro-inflammatory mediators.[41-45]
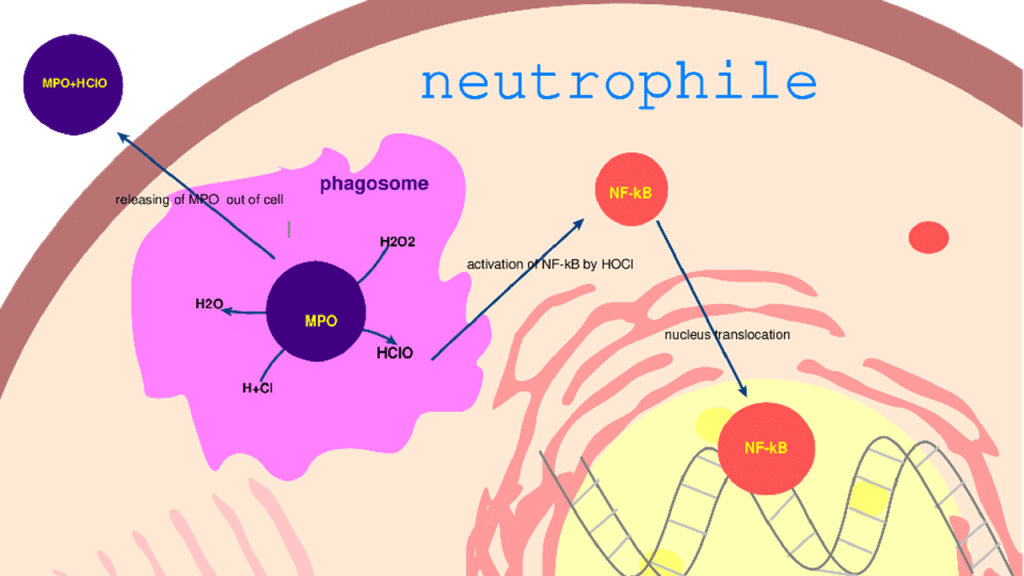
Figure 1: Mechanism of action of myeloperoxidase with production of reactive oxygen and nitrogen species
NADPH oxidase: It is an enzyme made up of 2 membrane subunits (Gp91phox and p22phox) and 4 cytosolic subunits (p40phox, p47phox, p67phox, and Rac).[46] It is present in neutrophils, and its production is correlated with the level of neutrophils present in the intestinal lumen (Table 2). The activation of this enzyme is via different signaling pathways through tyrosine kinase receptors, growth factor receptors or G protein-coupled receptors. The 2 main activation pathways of this enzyme are activation by phosphorylation of membrane receptors, protein kinase C (PKC), and activation by arachidonic acid and other fatty acids. It catalyzes the formation of the superoxide anion, which is a powerful oxidizing agent, thus causing the oxidative stress observed in these pathologies. It is the main enzyme of the respiratory outbreak (Figure 2).[47-50]
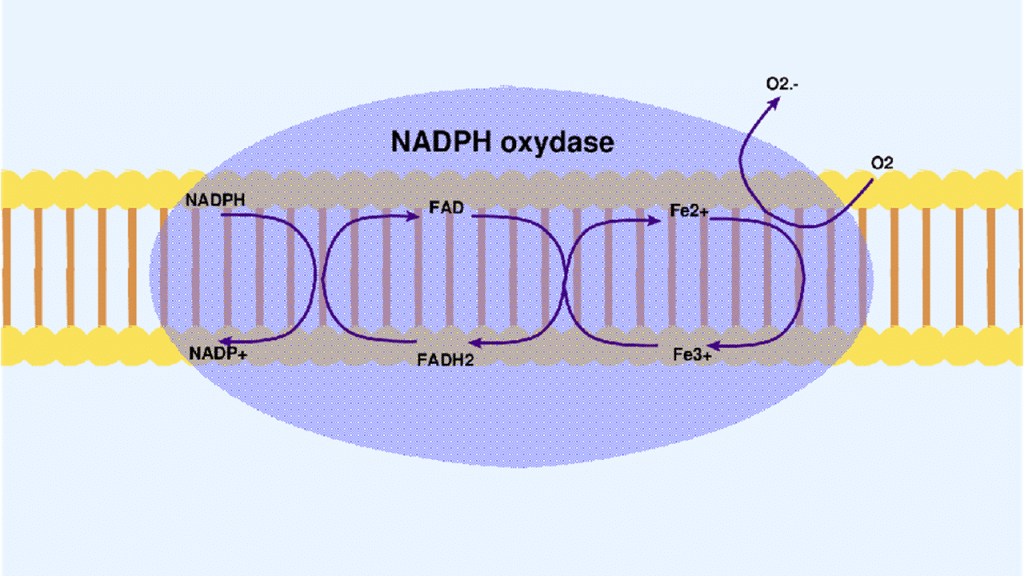
Figure 2: Mechanism of superoxide anion production by NADPH oxidase
Hemoxygenase-1: It is an enzyme that is a reference anti-inflammatory and antioxidant, and is expressed by various tissues in the body. HO-1 consists of 288 amino acid residues with its active site located between the first two alpha helices.[51] It converts heme to biliverdin, carbon monoxide (CO), and iron2+.[52] The CO produced inhibits the mitochondrial respiratory chain, thus preventing the phagocytic activity of macrophages. Similarly, the CO produced activates Tregs, which help to regulate the inflammatory response. The biliverdin produced decreases the expression of genes associated with cyclooxygenase-2, Nitric oxide synthase, prevents the infiltration of neutrophils into the wall of the jejunum, and decreases the production of IL-1β and IL-6 (Table 2). Its activation is under the control of several mediators such as heme, hemin, cobalt-protoporphyrin, and nrf-2 (Figure 3).[53-56]
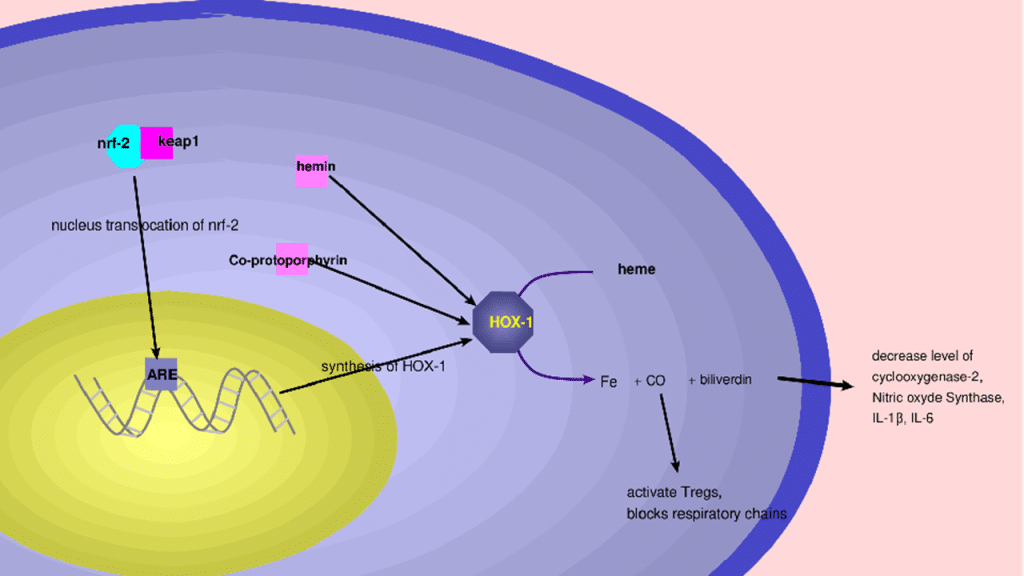
Figure 3: Hemoxygenase 1 activation mechanism by nrf2 and mechanism of action
Phosphodiesterase (PDE): PDE consists of a family of 7 isoenzymes that degrade cAMP and cGMP, which are implicated in the regulation of inflammation. Abnormal activation of PDE results in an abnormal response that leads to chronic inflammation of the intestinal wall. The main isoenzyme implicated in IBD is PDE4. PDE4 is a significant type of PDE family in immunocytes, which is responsible for degrading cyclic adenosine monophosphate (cAMP) in the cytoplasm. Some research reveals that high levels of cAMP in the cytoplasm can downregulate TNF-α generation in macrophages.[57-59] The use of PDE4 inhibitors increases cAMP and decreases the level of TNF-α as reported in research on the rat colon (Table 2). However, the main drugs that are actually used as PDE4 inhibitors show some inevitable adverse effects such as diarrhea, weight loss, nausea, and psychiatric events, which restrain the usage in UC management.[60-62]
Cyclooxygenase 2(COX-2): Cyclooxygenase consists of two isoforms, COX-1 and COX-2 that convert arachidonic acid to prostaglandins, protacyclins and thromboxans. These compounds play an important role in inflammation, such as vasodilatation to allow cell infiltration in inflamed tissues. COX-1 is produced under normal conditions, particularly in the endothelium, platelets, and kidneys, while COX-2 is inducible by inflammatory factors such as smoking, which is a risk factor in the etiology of Crohn’s disease (Table 2). COX-2 inhibitors are very interesting in IBD; however, they also block the production of PGI2, which is an important anti-aggregating agent.[63-68]
5-lipoxygenase(5-LOX): 5-LOX is an enzyme of a family of lipoxygenases that metabolize arachidonic acid into leukotrienes and lipoxins.[69] Certain Leukotriens like LTB4 play an important role in granulocytes and monocytes chemotaxis, recruitment, their degranulation with superoxide generation, increase cytokine production in LT cells, and increase permeability hyperalgesia, while LTC4 plays a role in intestinal contraction. All these mechanisms contribute in the development and maintain of IBD. Otherwise, lipoxins can be considered to have an antagonist role to leukotriens. They possess anti-inflammatory effects like LXA4 and LXB4, which reduce the production and chemotaxis of granulocytes and stimulate monocyte activation (Table 2). Lipoxins and synthetic lipoxin analogs inhibit transcription factors related to inflammatory processes like NF-κB, stimulate efferocytosis, reduce macrophage infiltration, and inhibit progression in some diseases such as atherosclerosis. All these make 5-lipoxygenase an interesting molecule in the regulation of IBD.[70-74]
Exosomes: They are small vesicles in biological fluids (urine, blood, milk, saliva, cerebrospinal fluid) formed from invaginations of lysosomes and which fuse with the cytoplasmic membrane before being released.[75] They are vesicles associated with intercellular communication through the transport of lipids, nucleic acids, and proteins (Figure 4).
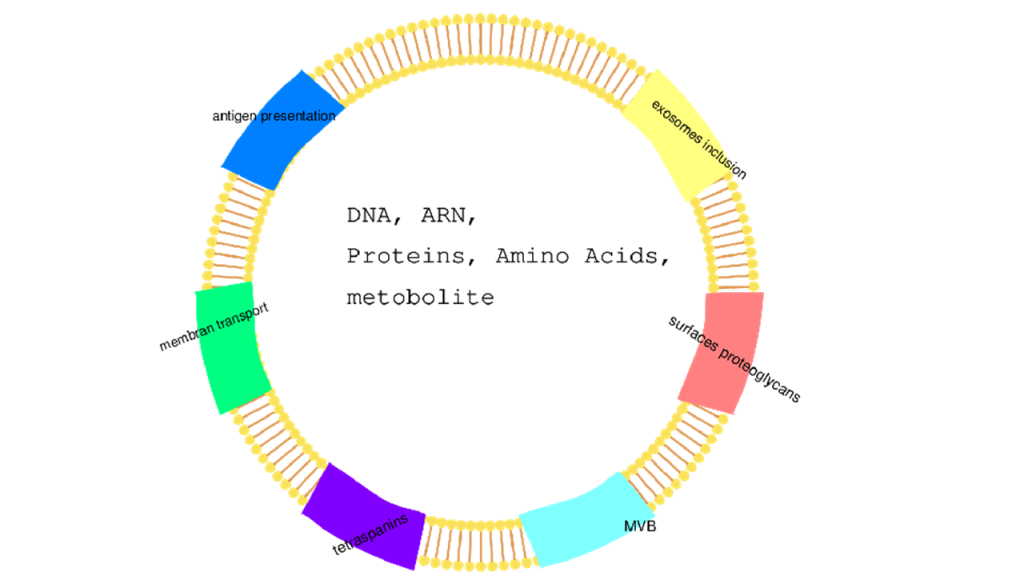
Figure 4: Exosome presentation
They express on their surface cellular markets such as α4β7 which is an integrin that is sympathetic to Vedolizumab which is an antibody directed against this same integrin on CD4+, thus decreasing the effectiveness of the treatment. Similarly, a study conducted on mitochondrial RNA of serum exosomes shows their implications on pro-inflammatory cytokine production and macrophage recruitment (Table 3).[76-78]
| Origins | Functions |
| body fluids and feces | novel biomarkers of IBD |
| human umbilical cord mesenchymal stem cells (hucMSCs);
colonic epithelial cells (CECs); mast cells (MCs) |
maintain the tight junction (TJ) barrier function |
| dendritic cells (DCs) | affect the intestinal microbiota profile in heat shock proteins-dependent manner |
| Intestinal epithelial cell (IEC)-derived | fuse with the DCs membrane to induce immune tolerance |
| colitis serum or treated DCs | regulate the immune cell proliferation through inflammation-related signaling pathways |
Table 3: Different types of exosomes and their functions
Transcription factors
Signal transducer and activator of transcription (STAT3): STAT3 is a member of the seven STAT transcription factors, which are present in an inactive state in the cytoplasm of the cells.[79,80] The binding of the specific cytokine to the cell surface receptor results in the activation of a receptor-associated Janus kinase (JAK) and leads to the phosphorylation of STAT3, initiating the pathway.[81-83] Each STAT3 has an SH2 domain (A phosphotyrosine recognition domain) through which it reciprocally interacts with the other tyrosine-phosphorylated STAT3 monomer, followed by translocation into the nucleus to transcribe the target genes.[84-86] STAT3 has been identified as an oncogenic transcription factor that has a major role in malignant transformation.[87,88] Constitutive activation of STAT3 signaling has been observed in several types of human cancers and cell lines.[89,90] The JAK-STAT pathway regulates the expression of multiple mediators involved in inflammatory pathways implicated in the pathogenesis of IBD.[91-94]
Nuclear factor kappa B (NF-kB): NF-κB is a complex of NF-κB and IκB-α in the cytoplasm. Inflammatory cytokines and intestinal microorganisms activate the NF-κB transcription factor process by inducing the phosphorylation and consequent degradation of IκB by its kinase.[95-99] This allows NF-κB translocation into the cell nucleus to activate gene expression for relevant inflammatory proteins which lead to the production of pro-inflammatory cytokines such as TNF-α, IL-12, IL-23, enzymes like COX-2, NADPH oxydase and iNOS, which are secreted from the cell and propagate the immune response by acting on neighboring immune cells. Additionally, activation of NF-κB is implicated in the oxidative stress process.[100-105]
Nuclear factor erythroid 2-related factor 2 (NRF-2): Nrf2 (nuclear factor erythroid 2-related factor 2) is a stress-responsive transcription factor, associated with cellular homeostasis, which is regulated by the bound of Keap1in cytosol. However, upon stimulation by electrophilic agents or compounds that possess the ability to modify thiol groups, the Keap1-mediated repression of Nrf2 activity is lost, allowing the translocation of Nrf2 to the nucleus and the potentiation of the ARE response to regulate the activities of target genes such as SOD, CAT, and those of phase II detoxifying enzymes to remove harmful molecules such as reactive oxygen species (ROS). The critical role of Keap1/NRF2 signaling is implicated in both acute and chronic inflammation of diseases beyond IBD, including rheumatoid arthritis, gastritis, asthma, and atherosclerosis (Figure 4).[106-110]
Sphingosine-1-phosphate (S1P): S1P is a lipid associated with immune cell dynamics and inflammation. S1P signaling on central memory T-cells allows their exit from lymph nodes. Internalization of the S1P receptor prevents lymphocytes from responding to S1P, so they can’t reach the inflamed tissue (Table 1). The level of S1P increases with the disruption of the mucosa in inflamed Tissue, which leads to the recruitment of immune cells and persistent inflammation and tissue damage. Drugs targeting S1P can modulate the progress of lymphocytes from the lymph nodes, reducing their availability to contribute to inflammation in the gut and help maintain the integrity of the intestinal barrier.[111-113]
Conclusion
IBD is a chronic disease of poorly understood etiology that remains a challenge for modern research. These currently incurable diseases are the focus of various fundamental and medical studies aimed at improving the health status of patients. The continuous search for new treatments is based on therapeutic targets defined as a priority in the evolution of these diseases. Neutrophils, macrophages, and lymphocytes are the main cellular targets maintaining this inflammatory state through the expression of various surface integrins, the production of lytic enzymes, and pro-inflammatory cytokines. Although research is more oriented towards anti-TNF-α, researchers are looking for new and more general therapeutic targets that may have multiple actions on the repression of these pathologies and thus improve the periods of remission of IBD.
References
- Moreno LO, Fernández-Tomé S, Abalo R. Biological treatments in inflammatory bowel disease: a complex mix of mechanisms and actions. Biologics. 2021;1(2):189-210. doi:10.3390/biologics1020012 Crossref | Google Scholar
- Benchimol EI, Bernstein CN, Bitton A, et al. The impact of inflammatory bowel disease in Canada 2018: a scientific report from the Canadian Gastro-Intestinal Epidemiology Consortium to Crohn’s and Colitis Canada. J Can Assoc Gastroenterol. 2019;2(Suppl 1):S1-5. doi:10.1093/jcag/gwy052 PubMed | Crossref | Google Scholar
- Ortiz T, Argüelles-Arias F, Illanes M, et al. Polyphenolic maqui extract as a potential nutraceutical to treat TNBS-induced Crohn’s disease by the regulation of antioxidant and anti-inflammatory pathways. Nutrients. 2020;12(6):1752. doi:10.3390/nu12061752 PubMed | Crossref | Google Scholar
- Krzesiek E, Kofla-Dlubacz A, Akutko K, Stawarski A. The incidence of inflammatory bowel disease in the paediatric population in the district of Lower Silesia, Poland. J Clin Med. 2021;10(17):3994. doi:10.3390/jcm10173994
PubMed | Crossref | Google Scholar - Hodges P, Kelly P. Inflammatory bowel disease in Africa: what is the current state of knowledge? Int Health. 2020;12(3):222-230. doi:10.1093/inthealth/ihaa005 PubMed | Crossref | Google Scholar
- Antunes JC, Seabra CL, Domingues JM, et al. Drug targeting of inflammatory bowel diseases by biomolecules. Nanomaterials. 2021;11(8):2035. doi:10.3390/nano11082035 PubMed | Crossref | Google Scholar
- Seyed Tabib NS, Madgwick M, Sudhakar P, et al. Big data in IBD: big progress for clinical practice. Gut. 2020;69(8):1520-1532. doi:10.1136/gutjnl-2019-320065 PubMed | Crossref | Google Scholar
- Torres J, Mehandru S, Colombel JF, Peyrin-Biroulet L. Crohn’s disease. Lancet. 2017;389(10080):1741-1755. doi:10.1016/S0140-6736(16)31711-1 PubMed | Crossref | Google Scholar
- Ungaro R, Mehandru S, Allen PB, Peyrin-Biroulet L, Colombel JF. Ulcerative colitis. Lancet. 2017;389(10080):1756-1770. doi:10.1016/S0140-6736(16)32126-2 PubMed | Crossref
- Annese V. Genetics and epigenetics of IBD. Pharmacol Res. 2020;159:104892. doi:10.1016/j.phrs.2020.104892
PubMed | Crossref | Google Scholar - Younis N, Zarif R, Mahfouz R. Inflammatory bowel disease: between genetics and microbiota. Mol Biol Rep. 2020;47(4):3053-3063. doi:10.1007/s11033-020-05318-5 PubMed | Crossref | Google Scholar
- Chougule P, Herlenius G, Hernandez NM, et al. Isolation and characterization of human primary enterocytes from small intestine using a novel method. Scand J Gastroenterol. 2012;47(11):1334-1343. doi:10.3109/00365521.2012.708940 PubMed | Crossref | Google Scholar
- Kmieć Z, Cyman M, Ślebioda TJ. Cells of the innate and adaptive immunity and their interactions in inflammatory bowel disease. Adv Med Sci. 2017;62(1):1-16. doi:10.1016/j.advms.2016.09.001 PubMed | Crossref | Google Scholar
- Abron JD, Singh NP, Price RL, et al. Genistein induces macrophage polarization and systemic cytokine to ameliorate experimental colitis. PLoS One. 2018;13(7):e0199631. doi:10.1371/journal.pone.0199631
PubMed | Crossref | Google Scholar - Seyedizade SS, Afshari K, Bayat S, et al. Current status of M1 and M2 macrophages pathway as drug targets for inflammatory bowel disease. Arch Immunol Ther Exp (Warsz). 2020;68(2):10. doi:10.1007/s00005-020-00576-4
PubMed | Crossref | Google Scholar - Lingxiao W, Hong L. Effects of resveratrol on the Nrf2 and HO-1 expression in diabetic vascular endothelial cells. Int J Clin Exp Med. 2017;10(1):684-691. Effects of resveratrol on the Nrf2 and HO-1 expression in diabetic vascular endothelial cells
- Sebastián VP, Salazar GA, Coronado-Arrázola I, et al. Heme oxygenase-1 as a modulator of intestinal inflammation development and progression. Front Immunol. 2018;9:1956. doi:10.3389/fimmu.2018.01956
PubMed | Crossref | Google Scholar - Binion DG, West GA, Ina K, et al. Enhanced leukocyte binding by intestinal microvascular endothelial cells in inflammatory bowel disease. Gastroenterology. 1997;112(6):1895-1907. doi:10.1053/gast.1997.v112.pm9178682
PubMed | Crossref | Google Scholar - Zundler S, Becker E, Weidinger C, Siegmund B. Anti-adhesion therapies in inflammatory bowel disease—molecular and clinical aspects. Front Immunol. 2017;8:891. doi:10.3389/fimmu.2017.00891 PubMed | Crossref | Google Scholar
- Park SC, Jeen YT. Anti-integrin therapy for inflammatory bowel disease. World J Gastroenterol. 2018;24(17):1868-1880. doi:10.3748/wjg.v24.i17.1868 PubMed | Crossref | Google Scholar
- Jang DI, Lee AH, Shin HY, et al. The role of tumor necrosis factor alpha (TNF-α) in autoimmune disease and current TNF-α inhibitors in therapeutics. Int J Mol Sci. 2021;22(5):2719. doi:10.3390/ijms22052719
PubMed | Crossref | Google Scholar - Friedrich M, Pohin M, Powrie F. Cytokine networks in the pathophysiology of inflammatory bowel disease. Immunity. 2019;50(4):992-1006. doi:10.1016/j.immuni.2019.03.017 PubMed | Crossref | Google Scholar
- Neurath MF. Cytokines in inflammatory bowel disease. Nat Rev Immunol. 2014;14(5):329-342. doi:10.1038/nri3661 PubMed | Crossref | Google Scholar
- Tracey D, Klareskog L, Sasso EH, Salfeld JG, Tak PP. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol Ther. 2008;117(2):244-279. doi:10.1016/j.pharmthera.2007.10.001
PubMed | Crossref | Google Scholar - Rehman M, Cancarevic I, Iskander B, Lalani S, Malik BH. Biologics targeting in the treatment of inflammatory bowel disease: a conundrum. Cureus. 2020;12(9):e10621. doi:10.7759/cureus.10621 PubMed | Crossref | Google Scholar
- Simon EG, Ghosh S, Iacucci M, Moran GW. Ustekinumab for the treatment of Crohn’s disease: can it find its niche? Therap Adv Gastroenterol. 2016;9(1):26-36. doi:10.1177/1756283X15618130 PubMed | Crossref | Google Scholar
- Benson JM, Peritt D, Scallon BJ, et al. Discovery and mechanism of ustekinumab: a human monoclonal antibody targeting interleukin-12 and interleukin-23 for treatment of immune-mediated disorders. MAbs. 2011;3(6):535-545. doi:10.4161/mabs.3.6.17815 PubMed | Crossref | Google Scholar
- Olivera P, Danese S, Peyrin-Biroulet L. JAK inhibition in inflammatory bowel disease. Expert Rev Clin Immunol. 2017;13(7):693-703. doi:10.1080/1744666X.2017.1291342 PubMed | Crossref | Google Scholar
- D’Amico F, Fiorino G, Furfaro F, Allocca M, Danese S. Janus kinase inhibitors for the treatment of inflammatory bowel diseases: developments from phase I and phase II clinical trials. Expert Opin Investig Drugs. 2018;27(7):595-599. doi:10.1080/13543784.2018.1492547 PubMed | Crossref | Google Scholar
- Musumeci F, Greco C, Giacchello I, et al. An update on JAK inhibitors. Curr Med Chem. 2019;26(10):1806-1832. doi:10.2174/0929867325666180327093502 PubMed | Crossref | Google Scholar
- Bechman K, Yates M, Galloway JB. The new entries in the therapeutic armamentarium: the small molecule JAK inhibitors. Pharmacol Res. 2019;147:104392. doi:10.1016/j.phrs.2019.104392 PubMed | Crossref | Google Scholar
- Virtanen AT, Haikarainen T, Raivola J, Silvennoinen O. Selective JAKinibs: prospects in inflammatory and autoimmune diseases. BioDrugs. 2019;33(1):15-32. doi:10.1007/s40259-019-00333-w PubMed | Crossref | Google Scholar
- Neurath MF. Current and emerging therapeutic targets for IBD. Gastroenterol Hepatol. 2017;14(11):688. doi:10.1038/nrgastro.2017.138 PubMed | Crossref | Google Scholar
- Klebanoff SJ. Myeloperoxidase: friend and foe. J Leukoc Biol. 2005;77(5):598-625. doi:10.1189/jlb.1204697
PubMed | Crossref | Google Scholar - Malle E, Furtmüller PG, Sattler W, Obinger C. Myeloperoxidase: a target for new drug development? Br J Pharmacol. 2007;152(6):838-854. doi:10.1038/sj.bjp.0707358 PubMed | Crossref | Google Scholar
- Cross AR, Segal AW. The NADPH oxidase of professional phagocytes–prototype of the NOX electron transport chain systems. Biochim Biophys Acta. 2004;1657(1):1-22. doi:10.1016/j.bbabio.2004.03.008
PubMed | Crossref | Google Scholar - Selemidis S, Sobey CG, Wingler K, Schmidt HH, Drummond GR. NADPH oxidases in the vasculature: molecular features, roles in disease and pharmacological inhibition. Pharmacol Ther. 2008;120(3):254-291. doi:10.1016/j.pharmthera.2008.08.005 PubMed | Crossref | Google Scholar
- Sheppard FR, Kelher MR, Moore EE, McLaughlin NJ, Banerjee A, Silliman CC. Structural organization of the neutrophil NADPH oxidase: phosphorylation and translocation during priming and activation. J Leukoc Biol. 2005;78(5):1025-1042. doi:10.1189/jlb.0804442 PubMed | Crossref | Google Scholar
- Ryter SW, Alam J, Choi AM. Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiol Rev. 2006;86(2):583-650. doi:10.1152/physrev.00011.2005 PubMed | Crossref | Google Scholar
- Loboda A, Damulewicz M, Pyza E, Jozkowicz A, Dulak J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: an evolutionarily conserved mechanism. Cell Mol Life Sci. 2016;73(17):3221-3247. doi:10.1007/s00018-016-2223-0 PubMed | Crossref | Google Scholar
- Riquelme SA, Pogu J, Anegon I, Bueno SM, Kalergis AM. Carbon monoxide impairs mitochondria-dependent endosomal maturation and antigen presentation in dendritic cells. Eur J Immunol. 2015;45(12):3269-3288. doi:10.1002/eji.201545671 PubMed | Crossref | Google Scholar
- Riquelme SA, Carreño LJ, Espinoza JA, et al. Modulation of antigen processing by haem-oxygenase 1. Implications on inflammation and tolerance. Immunology. 2016;149(1):1-12. doi:10.1111/imm.12605 PubMed | Crossref | Google Scholar
- Blancou P, Tardif V, Simon T, et al. Immunoregulatory properties of heme oxygenase-1. Methods Mol Biol. 2011;677:247-268. doi:10.1007/978-1-60761-869-0_18 PubMed | Crossref | Google Scholar
- Miao C, Wang X, Zhou W, Huang J. The emerging roles of exosomes in autoimmune diseases, with special emphasis on microRNAs in exosomes. Pharmacol Res. 2021;169:105680. doi:10.1016/j.phrs.2021.105680
PubMed | Crossref | Google Scholar - Ludwig N, Whiteside TL, Reichert TE. Challenges in exosome isolation and analysis in health and disease. Int J Mol Sci. 2019;20(19):4684. doi:10.3390/ijms20194684 PubMed | Crossref | Google Scholar
- Sandborn WJ, Baert F, Danese S, et al. Efficacy and safety of vedolizumab subcutaneous formulation in a randomized trial of patients with ulcerative colitis. Gastroenterology. 2020;158(3):562-572. doi:10.1053/j.gastro.2019.08.027 PubMed | Crossref | Google Scholar
- Piotrowska M, Swierczynski M, Fichna J, Piechota-Polanczyk A. The Nrf2 in the pathophysiology of the intestine: molecular mechanisms and therapeutic implications for inflammatory bowel diseases. Pharmacol Res. 2021;163:105243. doi:10.1016/j.phrs.2020.105243 PubMed | Crossref | Google Scholar
- El Miedany Y, Youssef S, Ahmed I, El Gaafary M. The gastrointestinal safety and effect on disease activity of etoricoxib, a selective COX-2 inhibitor in inflammatory bowel diseases. Am J Gastroenterol. 2006;101(2):311-317. doi:10.1111/j.1572-0241.2006.00384.x PubMed | Crossref | Google Scholar
- Geertsema S, Bourgonje AR, Fagundes RR, et al. The NRF2/Keap1 pathway as a therapeutic target in inflammatory bowel disease. Trends Mol Med. 2023;29(10):830-842. doi:10.1016/j.molmed.2023.07.008
PubMed | Crossref | Google Scholar - Martel-Pelletier J, Lajeunesse D, Reboul P, Pelletier JP. Therapeutic role of dual inhibitors of 5-LOX and COX, selective and non-selective non-steroidal anti-inflammatory drugs. Ann Rheum Dis. 2003;62(6):501-509. doi:10.1136/ard.62.6.501 PubMed | Crossref | Google Scholar
- Vavricka SR, Galván JA, Dawson H, et al. Expression patterns of TNFα, MAdCAM1, and STAT3 in intestinal and skin manifestations of inflammatory bowel disease. J Crohns Colitis. 2018;12(3):347-354. doi:10.1093/ecco-jcc/jjx158 PubMed | Crossref | Google Scholar
- Andersen V, Nimmo E, Krarup HB, et al. Cyclooxygenase-2 (COX-2) polymorphisms and risk of inflammatory bowel disease in a Scottish and Danish case-control study. Inflamm Bowel Dis. 2011;17(4):937-946. doi:10.1002/ibd.21440 PubMed | Crossref | Google Scholar
- Yeshi K, Jamtsho T, Wangchuk P. Current treatments, emerging therapeutics, and natural remedies for inflammatory bowel disease. Molecules. 2024;29(16):3954. doi:10.3390/molecules29163954 PubMed | Crossref | Google Scholar
- Rogler G. Efficacy of JAK inhibitors in Crohn’s disease. J Crohns Colitis. 2020;14(Supplement_2):S746-S754. doi:10.1093/ecco-jcc/jjz186 PubMed | Crossref | Google Scholar
- Chen D, Chen Y, Huang F, Zhang X, Zhou Y, Xu L. The underlying regulatory mechanisms of colorectal carcinoma by combining vitexin and aspirin: based on systems biology, molecular docking, molecular dynamics simulation, and in vitro study. Front Endocrinol (Lausanne). 2023;14:1147132. doi:10.3389/fendo.2023.1147132
PubMed | Crossref | Google Scholar - Lee JH, Mohan CD, Shanmugam MK, et al. Vitexin abrogates invasion and survival of hepatocellular carcinoma cells through targeting STAT3 signaling pathway. Biochimie. 2020;175:58-68. doi:10.1016/j.biochi.2020.05.006
PubMed | Crossref | Google Scholar - Massironi S, Furfaro F, Bencardino S, Allocca M, Danese S. Immunity in digestive diseases: new drugs for inflammatory bowel disease treatment-insights from phase II and III trials. J Gastroenterol. 2024;59(9):761-787. doi:10.1007/s00535-024-02130-x PubMed | Crossref | Google Scholar
- Swaminathan A, Borichevsky GM, Edwards TS, et al. Faecal myeloperoxidase as a biomarker of endoscopic activity in inflammatory bowel disease. J Crohns Colitis. 2022;16(12):1862-1873. doi:10.1093/ecco-jcc/jjac098
PubMed | Crossref | Google Scholar - Chen J, Xu F, Ruan X, et al. Therapeutic targets for inflammatory bowel disease: proteome-wide Mendelian randomization and colocalization analyses. EBioMedicine. 2023;89:104494. doi:10.1016/j.ebiom.2023.104494
PubMed | Crossref | Google Scholar - Cohen NA, Rubin DT. New targets in inflammatory bowel disease therapy: 2021. Curr Opin Gastroenterol. 2021;37(4):357-363. doi:10.1097/MOG.0000000000000740 PubMed | Crossref | Google Scholar
- Kumar A, Smith PJ. Horizon scanning: new and future therapies in the management of inflammatory bowel disease. BMJ eGastroenterol. 2023. Horizon scanning: new and future therapies in the management of inflammatory bowel disease
- AWARE-IBD Diagnostic Delay Working Group. Sources of diagnostic delay for people with Crohn’s disease and ulcerative colitis: Qualitative research study. PLoS One. 2024;19(6):e0301672. doi:10.1371/journal.pone.0301672
PubMed | Crossref | Google Scholar - Li X, Cole J, Vaughan D, Xiao Y, Walker D, Wall DM. Stratifying macrophages based on their infectious burden identifies novel host targets for intervention during Crohn’s disease associated adherent-invasive Escherichia coli infection. Microbiology. 2024;170(6):001470. doi:10.1099/mic.0.001470 PubMed | Crossref | Google Scholar
- Danese S, Furfaro F, Vetrano S. Targeting S1P in inflammatory bowel disease: new avenues for modulating intestinal leukocyte migration. J Crohns Colitis. 2018;12(suppl_2):S678-S686. doi:10.1093/ecco-jcc/jjx107
PubMed | Crossref | Google Scholar - Khan AA, Alsahli MA, Rahmani AH. Myeloperoxidase as an active disease biomarker: recent biochemical and pathological perspectives. Med Sci (Basel). 2018;6(2):33. doi:10.3390/medsci6020033 PubMed | Crossref | Google Scholar
- Diez-Martin E, Hernandez-Suarez L, Muñoz-Villafranca C, et al. Inflammatory bowel disease: a comprehensive analysis of molecular bases, predictive biomarkers, diagnostic methods, and therapeutic options. Int J Mol Sci. 2024;25(13):7062. doi:10.3390/ijms25137062 PubMed | Crossref | Google Scholar
- Núñez P, Quera R, Yarur AJ. Safety of Janus kinase inhibitors in inflammatory bowel diseases. Drugs. 2023;83(4):299-314. doi:10.1007/s40265-023-01840-5 PubMed | Crossref | Google Scholar
- Rogler G, Singh A, Kavanaugh A, Rubin DT. Extraintestinal manifestations of inflammatory bowel disease: current concepts, treatment, and implications for disease management. Gastroenterology. 2021;161(4):1118-1132. doi:10.1053/j.gastro.2021.07.042 PubMed | Crossref | Google Scholar
- Hamilton MJ, Snapper SB, Blumberg RS. Update on biologic pathways in inflammatory bowel disease and their therapeutic relevance. J Gastroenterol. 2012;47(1):1-8. doi:10.1007/s00535-011-0521-8 PubMed | Crossref | Google Scholar
- Piotrowska M, Binienda A, Fichna J. The role of fatty acids in Crohn’s disease pathophysiology – an overview. Mol Cell Endocrinol. 2021;538:111448. doi:10.1016/j.mce.2021.111448 PubMed | Crossref | Google Scholar
- O’Neill GP, Ford-Hutchinson AW. Expression of mRNA for cyclooxygenase-1 and cyclooxygenase-2 in human tissues. FEBS Lett. 1993;330(2):156-160. doi:10.1016/0014-5793(93)80263-t PubMed | Crossref | Google Scholar
- Felder JB, Korelitz BI, Rajapakse R, Schwarz S, Horatagis AP, Gleim G. Effects of nonsteroidal anti-inflammatory drugs on inflammatory bowel disease: a case-control study. Am J Gastroenterol. 2000;95(8):1949-1954. doi:10.1111/j.1572-0241.2000.02262.x PubMed | Crossref | Google Scholar
- de Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. 2016;13(1):13-27. doi:10.1038/nrgastro.2015.186 PubMed | Crossref | Google Scholar
- Chang JT. Pathophysiology of inflammatory bowel diseases. N Engl J Med. 2020;383(27):2652-2664. doi:10.1056/NEJMra2002697 PubMed | Crossref | Google Scholar
- Neurath MF. Current and emerging therapeutic targets for IBD. Nat Rev Gastroenterol Hepatol. 2017;14(5):269-278. doi:10.1038/nrgastro.2016.208 PubMed | Crossref | Google Scholar
- Shivaji UN, Nardone OM, Cannatelli R, Smith SC, Ghosh S, Iacucci M. Small molecule oral targeted therapies in ulcerative colitis. Lancet Gastroenterol Hepatol. 2020;5(9):850-861. doi:10.1016/S2468-1253(19)30414-5
PubMed | Crossref | Google Scholar - Baillie GS, Tejeda GS, Kelly MP. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: inhibition and beyond. Nat Rev Drug Discov. 2019;18(10):770-796. doi:10.1038/s41573-019-0033-4 PubMed | Crossref | Google Scholar
- Maurice DH, Ke H, Ahmad F, Wang Y, Chung J, Manganiello VC. Advances in targeting cyclic nucleotide phosphodiesterases. Nat Rev Drug Discov. 2014;13(4):290-314. doi:10.1038/nrd4228 PubMed | Crossref | Google Scholar
- Hatzelmann A, Schudt C. Anti-inflammatory and immunomodulatory potential of the novel PDE4 inhibitor roflumilast in vitro. J Pharmacol Exp Ther. 2001;297(1):267-279. Anti-inflammatory and immunomodulatory potential of the novel PDE4 inhibitor roflumilast in vitro
- Park DD, Yum HW, Zhong X, et al. Perilla frutescens extracts protect against dextran sulfate sodium-induced murine colitis: NF-κB, STAT3, and Nrf2 as putative targets. Front Pharmacol. 2017;8:482. doi:10.3389/fphar.2017.00482
PubMed | Crossref | Google Scholar - Ma Q. Role of Nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol. 2013;53:401-426. doi:10.1146/annurev-pharmtox-011112-140320 PubMed | Crossref | Google Scholar
- McMahon M, Itoh K, Yamamoto M, et al. The Cap’n’Collar basic leucine zipper transcription factor Nrf2 (NF-E2 p45-related factor 2) controls both constitutive and inducible expression of intestinal detoxification and glutathione biosynthetic enzymes. Cancer Res. 2001;61(8):3299-3307. The Cap’n’Collar basic leucine zipper transcription factor Nrf2 (NF-E2 p45-related factor 2) controls both constitutive and inducible expression of intestinal detoxification and glutathione biosynthetic enzymes
- Reginster JY, Bias P, Buchner A. First clinical results of Licofelone (ML 3000), an inhibitor of COX-1, COX-2, and 5-LOX, for the treatment of osteoarthritis. Ann Rheum Dis. 2002;61(suppl 1):189. First clinical results of Licofelone (ML 3000), an inhibitor of COX-1, COX-2, and 5-LOX, for the treatment of osteoarthritis
- Bray MA, Ford-Hutchinson AW, Smith MJ. Leukotriene B4: an inflammatory mediator in vivo. Prostaglandins. 1981;22(2):213-222. doi:10.1016/0090-6980(81)90036-8 PubMed | Crossref | Google Scholar
- Liu J, Di B, Xu LL. Recent advances in the treatment of IBD: Targets, mechanisms and related therapies. Cytokine Growth Factor Rev. 2023;71-72:1-12. doi:10.1016/j.cytogfr.2023.07.001 PubMed | Crossref | Google Scholar
- Hussenbux A, Silva A. Steroids in inflammatory bowel disease: a clinical review. J Prescribing Pract. 2021;3(3):107-111. doi:10.12968/jprp.2021.3.3.107 Crossref | Google Scholar
- Kaufmann HJ, Taubin HL. Nonsteroidal anti-inflammatory drugs activate quiescent inflammatory bowel disease. Ann Intern Med. 1987;107(4):513-516. doi:10.7326/0003-4819-107-4-513 PubMed | Crossref | Google Scholar
- Li H, Fan C, Feng C, et al. Inhibition of phosphodiesterase-4 attenuates murine ulcerative colitis through interference with mucosal immunity. Br J Pharmacol. 2019;176(13):2209-2226. doi:10.1111/bph.14667
PubMed | Crossref | Google Scholar - El-Ashmawy NE, Khedr NF, El-Bahrawy HA, El-Adawy SA. Roflumilast, type 4 phosphodiesterase inhibitor, attenuates inflammation in rats with ulcerative colitis via down-regulation of iNOS and elevation of cAMP. Int Immunopharmacol. 2018;56:36-42. doi:10.1016/j.intimp.2018.01.004 PubMed | Crossref | Google Scholar
- Li H, Zhang Y, Liu M, et al. Targeting PDE4 as a promising therapeutic strategy in chronic ulcerative colitis through modulating mucosal homeostasis. Acta Pharm Sin B. 2022;12(1):228-245. doi:10.1016/j.apsb.2021.04.007
PubMed | Crossref | Google Scholar - Lee JH, Kim C, Lee J, et al. Arctiin is a pharmacological inhibitor of STAT3 phosphorylation at tyrosine 705 residue and potentiates bortezomib-induced apoptotic and anti-angiogenic effects in human multiple myeloma cells. Phytomedicine. 2019;55:282-292. doi:10.1016/j.phymed.2018.06.038 PubMed | Crossref | Google Scholar
- Lee M, Hirpara JL, Eu JQ, et al. Targeting STAT3 and oxidative phosphorylation in oncogene-addicted tumors. Redox Biol. 2019;25:101073. doi:10.1016/j.redox.2018.101073 PubMed | Crossref | Google Scholar
- Lee JH, Kim C, Ko JH, et al. Casticin inhibits growth and enhances ionizing radiation-induced apoptosis through the suppression of STAT3 signaling cascade. J Cell Biochem. 2019;120(6):9787-9798. doi:10.1002/jcb.28259
PubMed | Crossref | Google Scholar - Lee JH, Kim C, Lee SG, et al. Ophiopogonin D, a steroidal glycoside abrogates STAT3 signaling cascade and exhibits anti-cancer activity by causing GSH/GSSG imbalance in lung carcinoma. Cancers (Basel). 2018;10(11):427. doi:10.3390/cancers10110427 PubMed | Crossref | Google Scholar
- Siveen KS, Sikka S, Surana R, et al. Targeting the STAT3 signaling pathway in cancer: role of synthetic and natural inhibitors. Biochim Biophys Acta. 2014;1845(2):136-154. doi:10.1016/j.bbcan.2013.12.005
PubMed | Crossref | Google Scholar - Johnston PA, Grandis JR. STAT3 signaling: anticancer strategies and challenges. Mol Interv. 2011;11(1):18-26. doi:10.1124/mi.11.1.4 PubMed | Crossref | Google Scholar
- Wong ALA, Hirpara JL, Pervaiz S, et al. Do STAT3 inhibitors have potential in the future for cancer therapy?. Expert Opin Investig Drugs. 2017;26(8):883-887. doi:10.1080/13543784.2017.1351941 PubMed | Crossref | Google Scholar
- Lee JH, Kim C, Baek SH, et al. Capsazepine inhibits JAK/STAT3 signaling, tumor growth, and cell survival in prostate cancer. Oncotarget. 2017;8(11):17700-17711. doi:10.18632/oncotarget.10775 PubMed | Crossref | Google Scholar
- Arora L, Kumar AP, Arfuso F, et al. The Role of Signal Transducer and Activator of Transcription 3 (STAT3) and Its Targeted Inhibition in Hematological Malignancies. Cancers (Basel). 2018;10(9):327. doi:10.3390/cancers10090327 PubMed | Crossref | Google Scholar
- Jung YY, Lee JH, Nam D, et al. Anti-myeloma effects of icariin are mediated through the attenuation of JAK/STAT3-dependent signaling cascade. Front Pharmacol. 2018;9:531. doi:10.3389/fphar.2018.00531
PubMed | Crossref | Google Scholar - Burke WM, Jin X, Lin HJ, et al. Inhibition of constitutively active Stat3 suppresses growth of human ovarian and breast cancer cells. Oncogene. 2001;20(55):7925-7934. doi:10.1038/sj.onc.1204990 PubMed | Crossref | Google Scholar
- Loh CY, Arya A, Naema AF, Wong WF, Sethi G, Looi CY. Signal Transducer and Activator of Transcription (STATs) Proteins in Cancer and Inflammation: Functions and Therapeutic Implication. Front Oncol. 2019;9:48. doi:10.3389/fonc.2019.00048 PubMed | Crossref | Google Scholar
- Mohan CD, Rangappa S, Preetham HD, et al. Targeting STAT3 signaling pathway in cancer by agents derived from Mother Nature. Semin Cancer Biol. 2022;80:157-182. doi:10.1016/j.semcancer.2020.03.016
PubMed | Crossref | Google Scholar - Scott O, Roifman CM. NF-κB pathway and the Goldilocks principle: Lessons from human disorders of immunity and inflammation. J Allergy Clin Immunol. 2019;143(5):1688-1701. doi:10.1016/j.jaci.2019.03.016
PubMed | Crossref | Google Scholar - Cui L, Wang W, Luo Y, et al. Polysaccharide from Scutellaria baicalensis Georgi ameliorates colitis via suppressing NF-κB signaling and NLRP3 inflammasome activation. Int J Biol Macromol. 2019;132:393-405. doi:10.1016/j.ijbiomac.2019.03.230 PubMed | Crossref | Google Scholar
- Korytina GF, Akhmadishina LZ, Aznabaeva YG, et al. Associations of the NRF2/KEAP1 pathway and antioxidant defense gene polymorphisms with chronic obstructive pulmonary disease. Gene. 2019;692:102-112. doi:10.1016/j.gene.2018.12.061 PubMed | Crossref | Google Scholar
- Dinkova-Kostova AT, Massiah MA, Bozak RE, Hicks RJ, Talalay P. Potency of Michael reaction acceptors as inducers of enzymes that protect against carcinogenesis depends on their reactivity with sulfhydryl groups. Proc Natl Acad Sci U S A. 2001;98(6):3404-3409. doi:10.1073/pnas.051632198 PubMed | Crossref | Google Scholar
- Martín-Montañez E, Pavia J, Valverde N, et al. The S1P mimetic fingolimod phosphate regulates mitochondrial oxidative stress in neuronal cells. Free Radic Biol Med. 2019;137:116-130. doi:10.1016/j.freeradbiomed.2019.04.022
PubMed | Crossref | Google Scholar - Khor TO, Huang MT, Kwon KH, Chan JY, Reddy BS, Kong AN. Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 2006;66(24):11580-11584. doi:10.1158/0008-5472.CAN-06-3562 PubMed | Crossref | Google Scholar
- Osburn WO, Karim B, Dolan PM, et al. Increased colonic inflammatory injury and formation of aberrant crypt foci in Nrf2-deficient mice upon dextran sulfate treatment. Int J Cancer. 2007;121(9):1883-1891. doi:10.1002/ijc.22943
PubMed | Crossref | Google Scholar - Qu SL, Chen L, Wen XS, et al. Suppression of Th17 cell differentiation via sphingosine-1-phosphate receptor 2 by cinnamaldehyde can ameliorate ulcerative colitis. Biomed Pharmacother. 2021;134:111116. doi:10.1016/j.biopha.2020.111116 PubMed | Crossref | Google Scholar
- Takabe K, Paugh SW, Milstien S, Spiegel S. “Inside-out” signaling of sphingosine-1-phosphate: therapeutic targets. Pharmacol Rev. 2008;60(2):181-195. doi:10.1124/pr.107.07113 PubMed | Crossref | Google Scholar
- Danese S, Furfaro F, Vetrano S. Targeting S1P in inflammatory bowel disease: new avenues for modulating intestinal leukocyte migration. J Crohns Colitis. 2018;12(suppl_2):S678-686. doi:10.1093/ecco-jcc/jjx107
PubMed | Crossref | Google Scholar
Acknowledgments
Not reported
Funding
None
Author Information
Corresponding Author:
Pechangou Nsangou Sylvain
Department of Biochemistry
University of Yaounde, Cameroon
Email: [email protected]
Co-Authors:
Ngohoba Vigny Sayal, Njayou Frédéric Nico, Moundipa Fewou Paul
Department of Biochemistry
University of Yaounde, Cameroon
Authors Contributions
All authors contributed to the conceptualization, investigation, and data curation by acquiring and critically reviewing the selected articles. They were collectively involved in the writing – original draft preparation, and writing – review & editing to refine the manuscript. Additionally, all authors participated in the supervision of the work, ensuring accuracy and completeness. The final manuscript was approved by all named authors for submission to the journal.
Ethical Approval
This is a review study that does not involve human or animal subjects. Hence not applicable.
Conflict of Interest Statement
The author declares no conflict of interest.
Guarantor
None
DOI
Cite this Article
Ngohoba VS, Pechangou NS, Njayou FN, Moundipa FP. Therapeutic Targets Associated with Medical Research in the Treatment of Inflammatory Bowel Disease. medtigo J Pharmacol. 2025;1(1):e3061214. doi:10.63096/medtigo3061214 Crossref













